Slice of PLOS: 57 Varieties of Tomato


This beautiful berry, domesticated in the Americas more than 2500 years ago, and introduced to the Old World in the 16th century, nowadays forms the basis of a $60bn worldwide industry. Although first treated with the suspicion that it might share the toxicity of its distant cousin the deadly nightshade, the fruit is now eaten raw, sundried, cooked in stews and sauces, or turned into ketchup. We borrowed its English name from the Aztecs (along with “chocolate”), but 500 years later Brits and Americans still can’t agree how to pronounce it (“Tomayto, tomahto? Let’s call the whole thing off”). Although the tomato owes its Linnaean name, Solanum lycopersicum, to Medieval German werewolf myths (the species name means “wolf peach”), some very modern science is being used to study its evolution, manipulate its properties, understand its biology and perfect its agriculture.
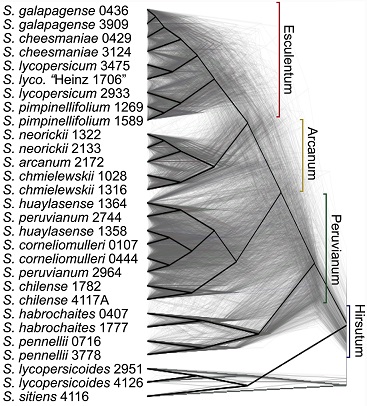
Artificial selection by humans has shaped tomatoes for the last few thousand years, but what happened before that? A recent tour de force by Leonie Moyle and colleagues, published in PLOS Biology, looks back over the 2,500,000 years that separate the existing 13 wild and domestic tomato species from their last common ancestor. Assembling transcriptome sequences from 29 plant varieties collected from the west coast of South America, the authors set out to use tomatoes as a case study in how evolution drives the emergence of diverse species. They see evidence that this burgeoning of diversity was fueled by three main sources of variation – cherry-picking of variation that was already present in the ancestral population, new mutations arising in the young species (including variants responsible for the red colour), and the shuffling of variants by interbreeding between species.

Although modern breeding practices have depleted some of the tomatoes’ ancestral diversity, a PLOS ONE paper shows that geographically localised European varieties (“landraces”) that have accumulated over the last 500 years still contain plenty of genetic variation that could be exploited to generate different shapes of tomatoes. But we no longer need to limit ourselves to spontaneously occurring mutations; a paper in Genome Biology from Daniel Voytas’ lab presents a system for targeted editing of the tomato genome. The authors use Agrobacterium to introduce bean yellow dwarf virus replicons containing targeted nucleases (either TALENs or CRISPR/Cas9) plus the sequence of interest. In this test case, they efficiently insert a strong promoter upstream of the ANT1 gene, successfully ramping up anthocyanin production and turning the entire tomato plant a deep purple.

Unsurprisingly, a substantial amount of tomato research is focused on understanding how they turn red. Two recent papers in PLOS Genetics by overlapping authors from Mondher Bouzayen and Mohamed Zouine’s group in France and Asaph Aharoni’s group in Israel flag the importance of two auxin response factors, ARF2A and ARF2B, in fruit ripening. Fruit lacking one or both of these proteins fail to turn red, and together the studies hint at considerable complexity, with ARF2A/B helping to integrate signalling from at least three plant hormones – auxin, ethylene and abscisic acid. A role for the last of these is explored in a PLOS ONE paper from Tiejin Ying’s group in Hangzhou – here they find that abscisic acid upregulates genes involved in making carotenoids and flavonoids and degrading chlorophyll; important steps in turning a green tomato red.

Finally, some elegant research is aimed at automating aspects of tomato agriculture. This PLOS ONE paper, for example, describes the use of combined thermal and 3D imaging to spot diseased plants. But why stop there? A study in Sensors describes an image processing system that spots ripe tomatoes among the foliage, with a view to telling a robotic harvester what to pick.
Credit for Featured Image: Flickr user adactio
For more detailed reading:
[or see this associated PLOS Collection]
Pease JB, Haak DC, Hahn MW, Moyle LC (2016) Phylogenomics Reveals Three Sources of Adaptive Variation during a Rapid Radiation. PLoS Biol 14(2): e1002379. doi: 10.1371/journal.pbio.1002379
Sacco A, Ruggieri V, Parisi M, Festa G, Rigano MM, Picarella ME, et al. (2015) Exploring a Tomato Landraces Collection for Fruit-Related Traits by the Aid of a High-Throughput Genomic Platform. PLoS ONE 10(9): e0137139. doi: 10.1371/journal.pone.0137139
Cermak T, Baltes NJ, Cegan R, Zhang Y, Voytas DF (2015) High-frequency precise modification of the tomato genome. Genome Biology 16:232. doi: 10.1186/s13059-015-0796-9
Hao Y, Hu G, Breitel D, Liu M, Mila I, Frasse P, et al. (2015) Auxin Response Factor SlARF2 Is an Essential Component of the Regulatory Mechanism Controlling Fruit Ripening in Tomato. PLoS Genet 11(12): e1005649. doi: 10.1371/journal.pgen.1005649
Breitel DA, Chappell-Maor L, Meir S, Panizel I, Puig CP, Hao Y, et al. (2016) AUXIN RESPONSE FACTOR 2 Intersects Hormonal Signals in the Regulation of Tomato Fruit Ripening. PLoS Genet 12(3): e1005903. doi: 10.1371/journal.pgen.1005903
Mou W, Li D, Luo Z, Mao L, Ying T (2015) Transcriptomic Analysis Reveals Possible Influences of ABA on Secondary Metabolism of Pigments, Flavonoids and Antioxidants in Tomato Fruit during Ripening. PLoS ONE 10(6): e0129598. doi: 10.1371/journal.pone.0129598
Raza S-e-A, Prince G, Clarkson JP, Rajpoot NM (2015) Automatic Detection of Diseased Tomato Plants Using Thermal and Stereo Visible Light Images. PLoS ONE 10(4): e0123262. doi: 10.1371/journal.pone.0123262
Zhao Y, Gong L, Huang Y, Liu C (2016) Robust Tomato Recognition for Robotic Harvesting Using Feature Images Fusion. Sensors 16(2): 173. doi: 10.3390/s16020173