The XV Collection: Re-Interpreting Pheromone Signaling

by Piali Sengupta
Negative results that call into question a previously proposed model are generally difficult to publish unless they overturn a particularly high profile finding. And yet, negative results are critically important for the advancement of knowledge; if a model isn’t supported by further experiments it must be reconsidered.
For the PLOS Biology XV Collection, I’ve chosen to highlight a 2013 article by Gomez-Diaz et al., “Ligands for Pheromone-Sensing Neurons Are Not Conformationally Activated Odorant Binding Proteins” [1]. This article experimentally addresses a previously published model for how a pheromone signals via its receptor in the fruit fly Drosophila melanogaster [2]. Negative data from Gomez-Diaz and colleagues indicated that this proposed model is likely incorrect.
Pheromones are small molecules that mediate intraspecific chemical communication and convey information about an individual’s social status, health, and sex, among others. In the individual that receives the signal, pheromones elicit long-term changes in physiology and development, as well as rapid changes in behavior. Decoding the complex language of pheromone signaling remains an ongoing challenge.
Pheromones play a particularly critical role in the lifecycles of insects. In Drosophila, the male-specific pheromone cis-vaccenyl acetate (cVA) regulates courtship and aggression behaviors. This pheromone is sensed via the Or67d olfactory receptor, which expressed in a subset of olfactory sensory neurons (OSNs) located in the T1 trichoid sensilla on the fly’s antennae.

Odorant-binding proteins (OBPs) are diverse proteins that facilitate the presentation of odorants to their cognate receptors, but aren’t thought to directly bind the receptors themselves. cVA signaling via Or67d requires an OBP called LUSH. Previous work had shown that in the absence of LUSH, basal spiking of Or67d-expressing OSNs decreased, leading to the hypothesis that LUSH may directly bind Or67d and thereby modulate OSN firing.
Laughlin et al [2] experimentally tested this hypothesis by generating a presumed constitutively active recombinant LUSH protein (LUSHD118A). They then infused this recombinant protein into the sensilla of lush mutant flies by including the recombinant protein in the recording pipet. This protein was found to dramatically increase spontaneous firing of the Or67d neurons; activity was not further increased upon cVA addition. These data were interpreted to support the notion that LUSH acts directly on Or67d.
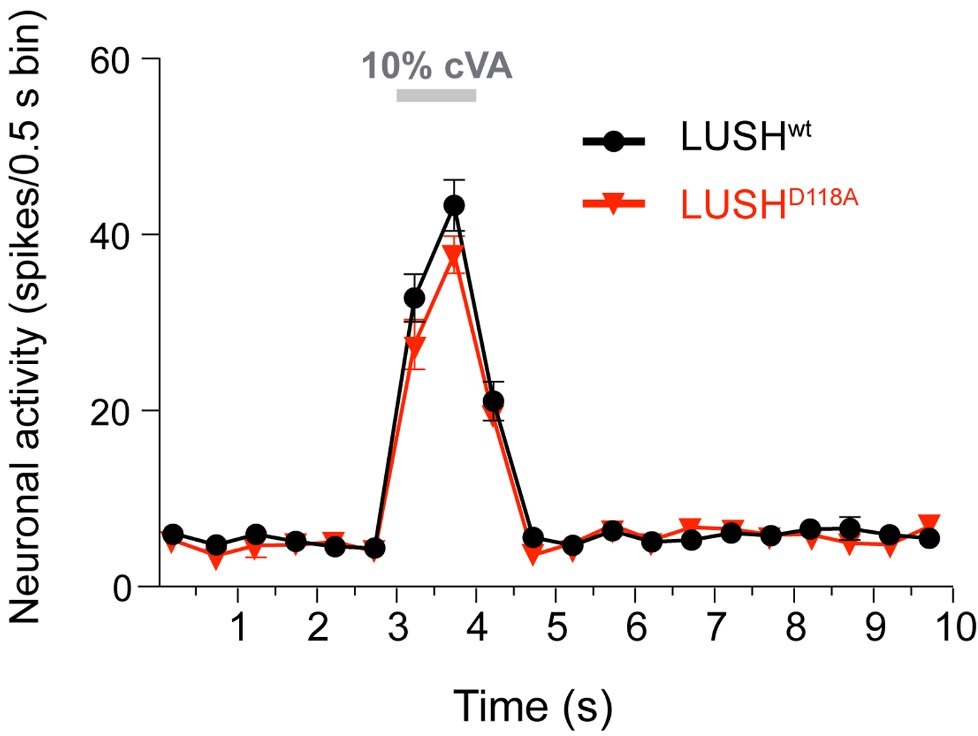
Gomez-Diaz and co-workers used a different experimental approach to test this model. They generated transgenic flies expressing LUSHD118A, the same presumed constitutively active mutant, at endogenous levels under the control of lush regulatory sequences. The transgenic LUSHD118A did not result in the enhanced OSN activity that Laughlin et al. had observed in the infusion experiment. Moreover, these neurons retained the ability to be activated by cVA, and lush mutant flies expressing additional LUSH mutant proteins at endogenous levels didn’t recapitulate the increased or decreased cVA sensitivity shown by Laughlin et al.
What could account for this discrepancy? Recombinant LUSH protein purified from bacteria may have different properties than endogenously produced protein. Acute presentation of LUSH proteins at non-physiological concentrations may also have elicited non-physiological effects. Indeed, an independent transgenic strain expressing LUSHD118A [3] also failed to recapitulate the infusion results.
The negative results reported by Gomez-Diaz and co-workers indicated that LUSH is unlikely to directly activate Or67d in the manner proposed in previous publications. This paper doesn’t include new experiments addressing LUSH function, and indeed the role of LUSH in pheromone signaling remains mysterious to this day. But by presenting these negative data, the work described by Gomez-Diaz et al is important as it rules out one potential hypothesis and allows the field to continue exploring exactly how pheromones collaborate with OBPs to elicit specific behaviors.
- Gomez-Diaz C, Reina JH, Cambillau C, Benton R. Ligands for pheromone-sensing neurons are not conformationally activated odorant binding proteins. PLoS Biol. 2013;11(4):e1001546. https://doi.org/10.1371/journal.pbio.1001546
- Laughlin JD, Ha TS, Jones DNM, Smith DP. Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell. 2008;133(7):1255-65. https://doi.org/10.1016/j.cell.2008.04.046
- Ronderos DS, Smith DP. Activation of the T1 neuronal circuit is necessary and sufficient to induce sexually dimorphic mating behavior in Drosophila melanogaster. J Neurosci. 2010;30(7):2595-9. https://doi.org/10.1523/JNEUROSCI.4819-09.2010
Piali Sengupta is at the Department of Biology and National Center for Behavioral Genomics, Brandeis University, and is a member of the PLOS Biology Editorial Board.

This blog post is the sixth in a series of twelve, forming PLOS Biology’s XV Collection, celebrating 15 years of outstanding open science; read Lauren Richardson’s blog for more information.
Featured image credit: pbio.1001546
Piali Sengupta image credit: Chad Nusbaum